 腾讯登录
腾讯登录代谢流终极科普:一文讲透细胞里的“交通网络”(建议收藏)
| 导读 | 绘云生物长期专注于医学代谢组学、多组学技术和稳定同位素代谢流平台建设。 |
一、为什么代谢流突然火了?从“代谢物存在”到“代谢物流动”
最近几年,肿瘤代谢、GLP-1 减肥药、CAR-T、长寿研究、线粒体医学、免疫代谢等方向迅速升温。看似完全不同的研究热点,背后其实都在关注同一个核心问题:
细胞里的物质与能量,究竟是如何流动的?
过去几十年,生命科学的发展一直围绕“细胞里有什么”展开。从基因组学、转录组学、蛋白组学到代谢组学,研究者不断尝试从不同层面解析生命系统的运行规律。其中,代谢组学之所以重要,是因为代谢距离细胞表型最近。细胞增殖、免疫激活、干细胞分化、肿瘤发生、衰老等生命活动,几乎都伴随着代谢网络的重编程。但随着研究深入,人们逐渐发现:传统代谢组学测到的,主要是代谢物浓度。
而浓度,并不等于通路活性。
例如,同样是肿瘤细胞乳酸升高,背后的机制可能完全不同:一种情况是糖酵解极度活跃,大量葡萄糖被快速转化为乳酸;另一种情况则可能是乳酸后续代谢受阻,导致其在细胞内堆积。虽然两种情况都会表现为乳酸浓度增加,但对应的代谢状态、疾病机制乃至治疗策略却截然不同。这就像城市道路中的汽车:路上车很多,既可能说明车流量巨大,也可能只是因为发生了堵车。传统代谢组学能够统计“有多少车”,却很难回答这些车从哪里来、正在往哪里去、哪条道路最繁忙、哪里才是真正的瓶颈。于是,生命科学开始从研究“代谢物存在”,逐渐转向研究“代谢物流动”。
随着稳定同位素示踪、质谱检测和数学建模的发展,代谢流分析(Metabolic Flux Analysis, MFA)逐渐成熟。它让研究者不再只看某一时刻的代谢物丰度,而是进一步追踪碳原子、氮原子等在代谢网络中的真实流向。代谢研究,也由此从“静态代谢物检测”,进入了“动态代谢网络解析”时代。
二、什么是代谢流?
代谢流(Metabolic Flux),本质上是指代谢网络中代谢物通过各条生化反应通路的实际速率与方向。简单来说,它研究的不是“细胞里有什么代谢物”,而是“这些代谢物究竟如何流动”。
传统代谢组学更像是在给细胞拍一张“静态照片”,能够看到某一时刻代谢物的丰度;而代谢流分析则更像是在拍摄“实时交通监控”,关注的是整个代谢网络中的动态变化过程。目前最常用的代谢流研究方法,是稳定同位素示踪技术。研究者会向细胞或动物体系中加入带有稳定同位素标记的代谢底物,例如:
* ¹³C₆-葡萄糖
* ¹³C₅-谷氨酰胺
* ¹⁵N-谷氨酸等
这些带有“标记”的碳原子进入细胞后,会沿着糖酵解、三羧酸循环(TCA)、磷酸戊糖途径以及脂肪酸合成等代谢通路不断转移、拆分与重组。不同代谢通路的活跃程度不同,就会形成不同的同位素标记模式。随后,研究者通过 LC-MS/MS 等技术检测代谢物的同位素分布情况,再结合数学模型进行反推,就能够计算出:哪条通路真正活跃;不同碳源的贡献比例;代谢流向如何重新分配;每一步反应的真实通量。
如果把细胞代谢系统比作城市交通网络:
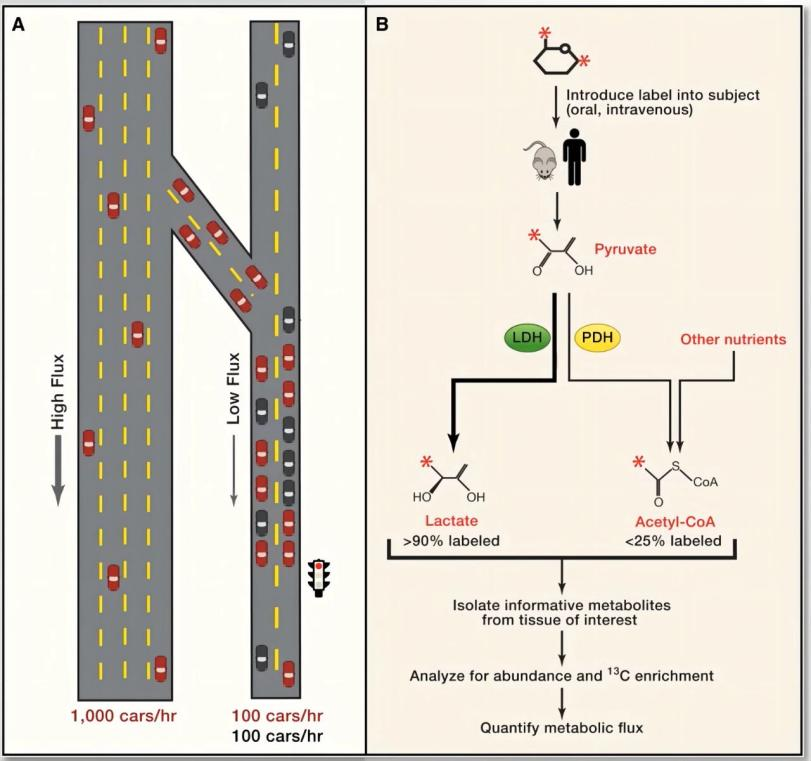
图源:DeBerardinis RJ, Thompson CB. Cell. 2012
传统代谢组学看到的是“车的数量”,而代谢流分析真正研究的则是:车从哪里进入城市、沿着哪条道路流动、哪里发生拥堵、哪些支路被重新激活。因此,代谢流分析研究的并不是静态的“代谢物库存”,而是动态的“代谢物流量”。它第一次让研究者能够真正“看见”细胞内部物质与能量的流动过程。
三、代谢流核心原理:同位素示踪
代谢流分析之所以能够“看见”代谢物流动,关键在于稳定同位素示踪。简单来说,就是给进入细胞的营养底物做上“标记”。研究者会向细胞或动物体系中加入带有稳定同位素标记的底物,例如 ¹³C₆-葡萄糖、¹³C₅-谷氨酰胺、¹⁵N 标记氨基酸等。进入细胞后,这些被标记的碳原子或氮原子会随着代谢反应不断转移,沿着糖酵解、TCA 循环、磷酸戊糖途径、脂肪酸合成、氨基酸代谢等通路发生拆分、重组和分流。
不同通路的活跃程度不同,最终会形成不同的同位素标记模式。通过 LC-MS/MS 检测这些标记模式,再结合天然同位素校正和数学模型分析,研究者就可以反推出碳源贡献、通路活性和代谢流向变化。
以丙酮酸为例,它含有 3 个碳原子。如果没有碳被 ¹³C 标记,就是 M+0;如果有 1 个碳被标记,就是 M+1;2 个碳被标记,就是 M+2;3 个碳全部被标记,就是 M+3。这些不同的 M+n 信号,就像代谢物在细胞中留下的“轨迹记录”。代谢流分析,正是通过读取这些轨迹,来判断代谢物从哪里来、流向哪里,以及哪条通路真正被激活。
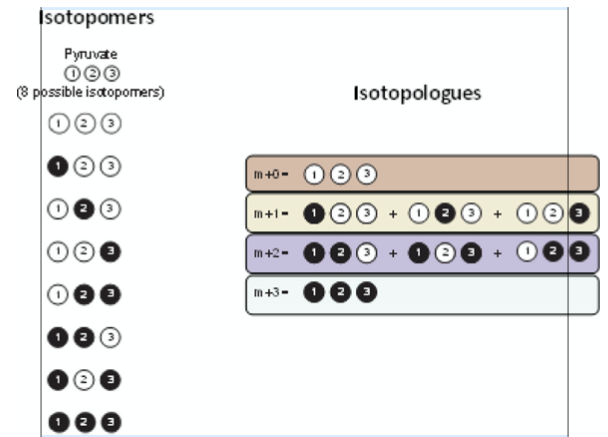
图源:Moiz B, et al. Trends in Endocrinology & Metabolism. 2023.
下图全面总结了常用的同位素示踪剂及其通路研究。
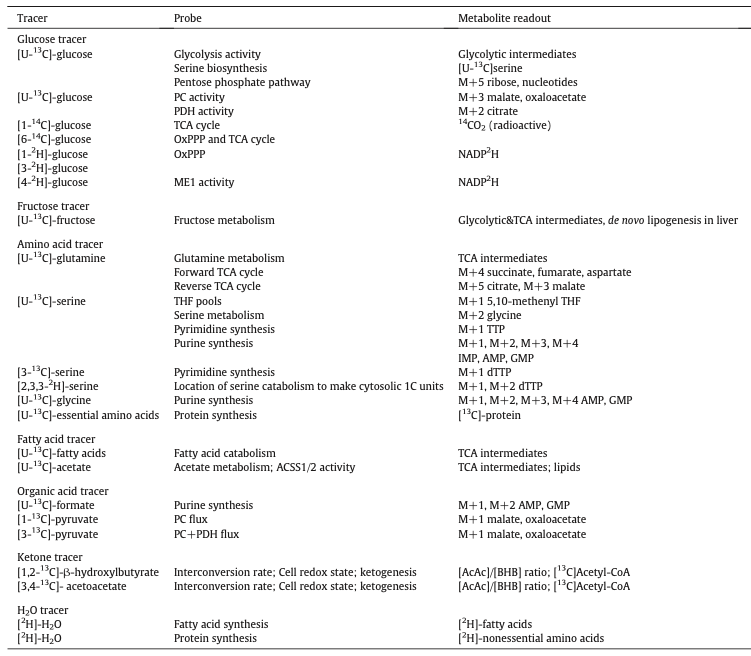
参考自:Liang L, et al.Pharmacology & Therapeutics. 2021.
四、代谢流实验流程:从科学问题到机制解析
代谢流实验并不是简单地“加一个同位素底物,然后上机检测”。它真正的起点,是一个明确的科学问题:研究者究竟想知道哪条代谢通路发生了变化?
代谢流实验最终要回答的是:
这些碳从哪里来?
进入了哪条通路?
在哪个节点发生分流?
哪条代谢路径被激活或抑制?
这些变化又如何解释具体的生物学机制?
因此,代谢流实验的完整逻辑可以概括为:
科学问题提出
首先明确实验真正想回答的问题,例如某条代谢通路是否被激活、某种营养底物是否进入特定合成途径,或某个代谢节点是否发生分流。
→ 示踪策略设计
根据研究问题选择合适的同位素标记底物、标记方式、实验体系和采样时间点,这是决定代谢流实验能否回答问题的关键。
→ 同位素标记实验
将带有 ¹³C、¹⁵N 或 ²H 等稳定同位素标记的底物加入细胞、动物或其他实验体系中,让标记原子进入真实的代谢网络。
→ 样本采集与代谢物提取
在设定时间点快速收集细胞、培养上清、血液或组织样本,并通过规范的前处理方法提取目标代谢物,尽量保留当时的代谢状态。
→ LC-MS/MS 检测
利用液相色谱-串联质谱检测代谢物及其不同同位素标记形式,获得 M+0、M+1、M+2 等同位素分布信息。
→ MID/MPE 数据分析
通过天然同位素丰度校正后,分析同位素分布模式和摩尔百分富集度,判断标记底物对不同代谢物的贡献比例。
→ 代谢流向解析
结合代谢通路知识和标记模式,推断碳源或氮源从哪里来、流向哪里,以及哪些通路或分支发生了增强、减弱或重新分配。
→ 机制阐释
最终将代谢流变化与具体生物学现象联系起来,解释疾病发生、药物作用、细胞命运改变或功能调控背后的代谢机制。
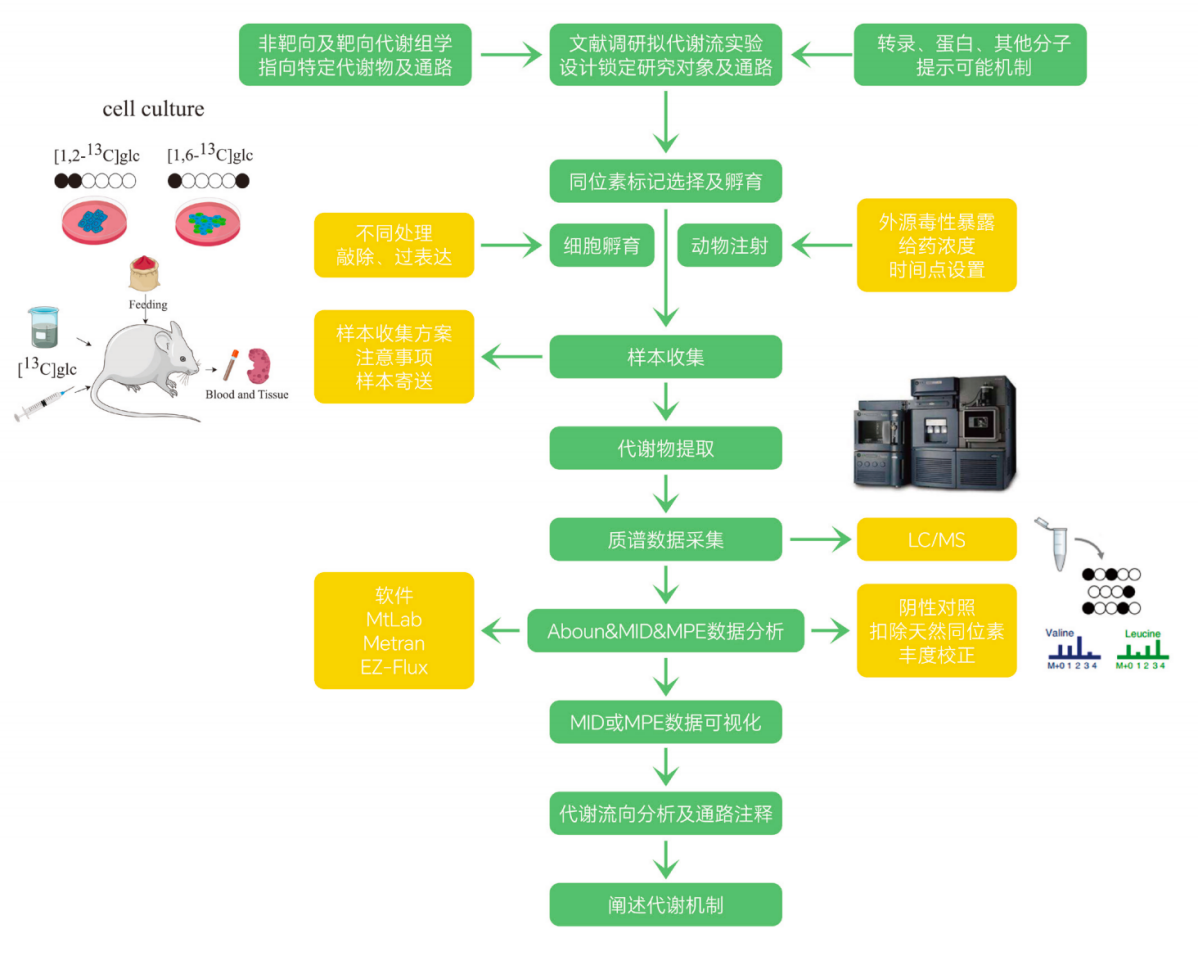
这也是代谢流分析区别于普通代谢组学检测的关键所在:
它不是单纯“测代谢物”,而是围绕一个明确的生物学问题,追踪代谢物在细胞或动物体内的真实流动过程。
五、数据分析结果:代谢流数据到底怎么看?
经过 LC-MS/MS 检测后,代谢流实验得到的并不是简单的“代谢物浓度表”,而是一组带有同位素标记信息的数据。它的核心价值在于:通过不同代谢物的标记模式,判断标记底物是否进入目标通路,以及不同代谢路径之间是否发生了流向变化。
在正式分析前,首先需要进行天然同位素丰度校正。因为自然界中的代谢物本身就含有一定比例的 ¹³C、²H、¹⁵N 等同位素,所以质谱检测到的 M+1、M+2 信号,并不全部来自实验加入的标记底物。只有扣除这部分天然本底信号后,才能得到更准确的同位素标记比例。
1. MID:看标记的“分布模式”
MID,也就是同位素分布模式,反映的是一个代谢物中 M+0、M+1、M+2、M+n 等不同标记形式所占的比例。它相当于代谢物的“同位素指纹”。
不同通路的活跃程度不同,最终会形成不同的 MID 模式。因此,MID 常用于判断标记碳原子是否进入了某条代谢通路,以及不同通路之间是否发生了流向变化。
2. MPE:看标记的“总体占比”
MPE,也就是摩尔百分富集度,用来表示某个代谢物中有多少比例的原子来自标记底物。
如果说 MID 更像是在看“标记分布长什么样”,那么 MPE 更关注的是:这个代谢物整体上有多少来自我们加入的示踪底物。因此,MPE 常用于评估某种营养底物对目标代谢物的贡献比例。
简单总结:
MID 看的是“标记分布”;
MPE 看的是“碳源贡献”。
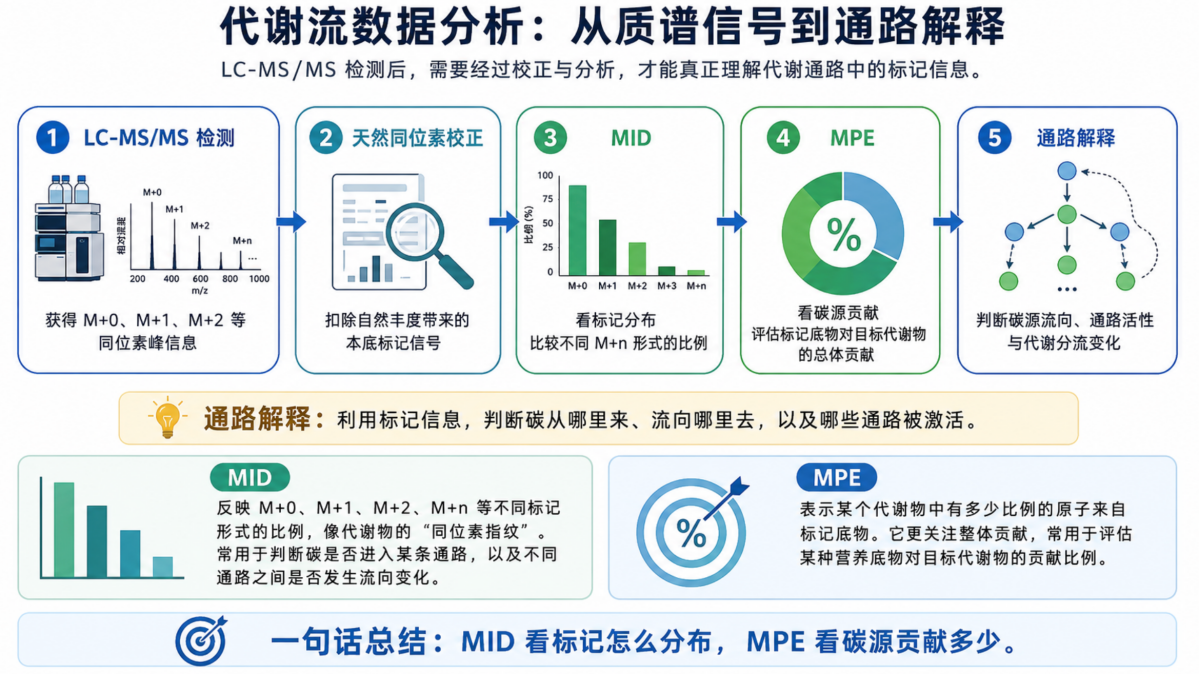
因此,代谢流数据分析的重点并不是单纯得到一组数值,而是通过这些标记信息回答:
碳从哪里来?
流向哪里去?
哪条通路被激活?
哪个节点发生了分流?
细胞的代谢网络如何被重新编程?
六、代谢流应用示例
示例一:同位素标记如何揭示“看不见”的代谢流向?
如果不使用同位素标记,我们通常只能得到比较有限的信息。例如,可以检测到葡萄糖被消耗,也可以看到某些糖酵解中间产物或下游产物发生变化。但这些数据很难告诉我们:
是否发生重新分配?
某些看似稳定的代谢物背后,是否存在活跃的循环流动?
这正是稳定同位素示踪的价值所在。
当研究者使用特定位点标记的葡萄糖,例如 [1,2-¹³C₂]-Glucose 进行示踪时,¹³C 标记会随着糖酵解反应进入下游代谢物。由于不同反应会导致碳骨架以不同方式拆分和重组,最终会在果糖-1,6-二磷酸、GAP 等代谢物中形成不同的同位素标记模式。
通过分析这些 M+0、M+2、M+4 等标记分布,研究者不仅可以判断碳是否进入了目标通路,还可以进一步发现某些隐藏的反应方向,例如糖酵解中的可逆反应和中间产物交换。
这说明,代谢流分析真正强大的地方在于:
它不仅能看到“代谢物变了”,还能看到代谢网络内部真实发生了什么。
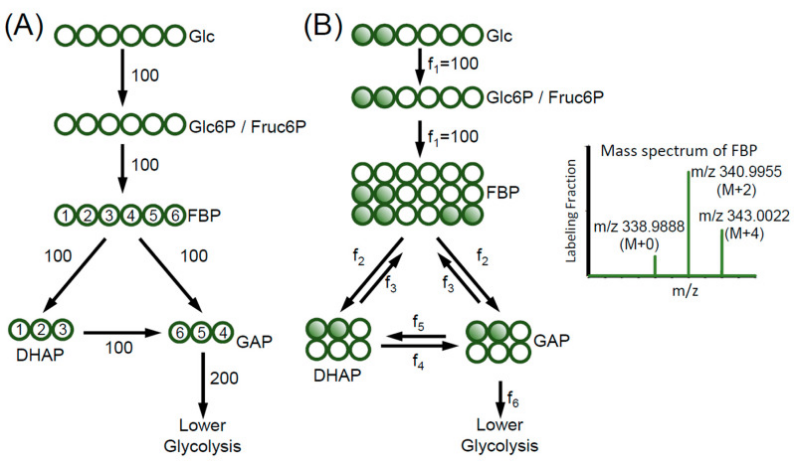
图源:Wang Y, et al. Metabolites. 2020.
示例二:[U-¹³C₆]-葡萄糖如何追踪 TCA 循环和脂肪酸合成?
[U-¹³C₆]-葡萄糖是代谢流研究中最常用的示踪底物之一。
它的 6 个碳原子全部被 ¹³C 标记,进入细胞后,可以沿着多条重要代谢通路流动。
首先,葡萄糖经过糖酵解生成丙酮酸;随后,丙酮酸可以转化为乳酸,也可以进入线粒体,通过 PDH 生成乙酰辅酶 A,进一步进入三羧酸循环(TCA)。
在 TCA 循环中,¹³C 标记会逐步进入柠檬酸、α-酮戊二酸、琥珀酸、延胡索酸、苹果酸等中间产物,形成不同的同位素分布模式。
这些模式可以反映:
* 葡萄糖碳是否进入 TCA 循环
* 氧化代谢是否增强
* 丙酮酸羧化酶等回补反应是否活跃
* TCA 中间产物是否发生重新分配
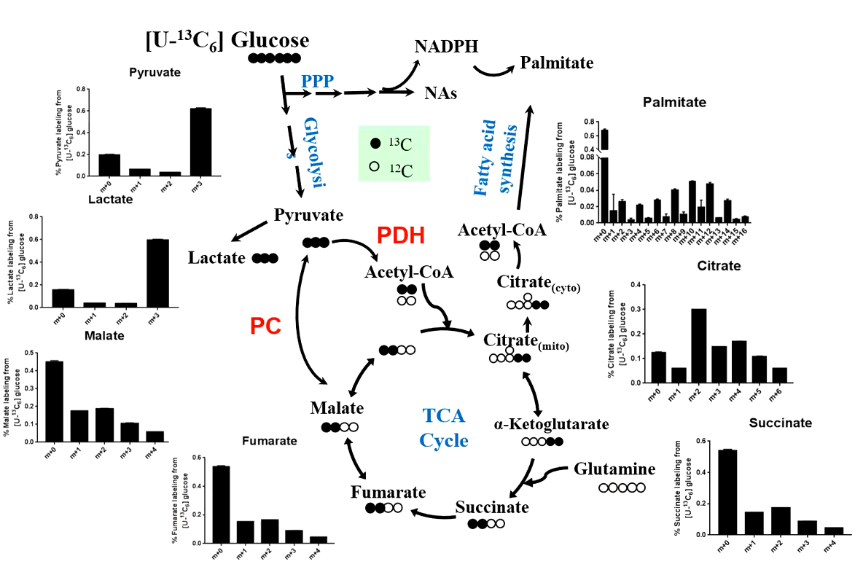
与此同时,葡萄糖还可以进入磷酸戊糖途径,产生 NADPH,为脂肪酸合成提供还原力;线粒体中的柠檬酸也可以转运到细胞质,裂解生成乙酰辅酶 A,进一步用于脂肪酸合成。
因此,通过追踪 ¹³C 从葡萄糖进入 TCA 循环和棕榈酸等脂质分子的过程,研究者可以同时评估细胞的氧化代谢和合成代谢活性。
简单来说:
[U-¹³C₆]-葡萄糖就像一条贯穿糖酵解、TCA 循环、磷酸戊糖途径和脂肪酸合成的“主线”。
通过观察这条主线上的标记分布,我们可以判断细胞究竟是在更多地燃烧葡萄糖,还是把葡萄糖用于合成新的生物大分子。
七、代谢流研究文献案例
前面两个示例展示了代谢流的基本分析思路。接下来,我们通过两项真实研究,看代谢流如何在疾病机制研究中发挥作用。
文献案例一:代谢流揭示脑缺血中的线粒体转移机制
文章标题:Astrocytic LRP1 enables mitochondria transfer to neurons and mitigates brain ischemic stroke by suppressing ARF1 lactylation
期刊:Cell Metabolism
发表时间:2024 年
影响因子:30.9
在脑缺血损伤中,神经元能量代谢受损是导致细胞死亡的重要原因。近年来,越来越多研究发现,星形胶质细胞不仅可以通过分泌因子保护神经元,还可能通过线粒体转移的方式,为受损神经元提供代谢支持。
但一个关键问题是:
这些线粒体转移背后,究竟受到怎样的代谢调控?
在《Cell Metabolism》发表的研究中,利用稳定同位素代谢流技术,结合 [U-¹³C₆]-Glucose、[U-¹³C₅]-Glutamine 和 [U-¹³C₂]-Acetate 等示踪策略,追踪星形胶质细胞中的糖酵解、乳酸代谢和 TCA 循环变化。研究发现,星形胶质细胞中的 LRP1 能够通过抑制 ARF1 乳酸化,促进线粒体向神经元转移,从而缓解缺血性脑卒中损伤。
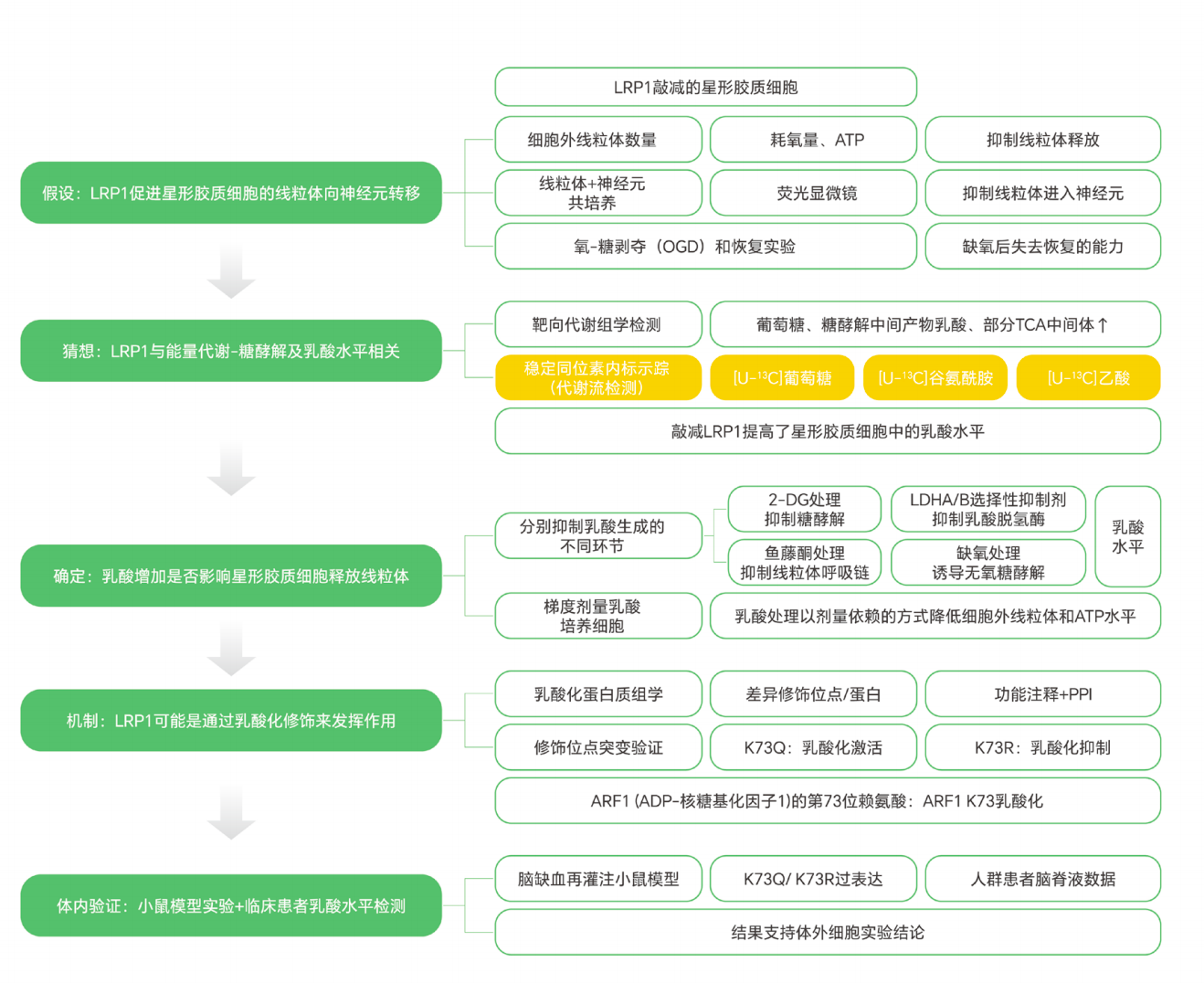
这个案例说明,代谢流分析已经不只是观察“某个代谢物是否升高”,而是可以进一步解析代谢重编程如何参与细胞互作、线粒体转移和疾病进展。
文献案例二:代谢流解析骨髓微环境中丝氨酸如何影响多发性骨髓瘤
文章标题:Excessive serine from the bone marrow microenvironment impairs megakaryopoiesis and thrombopoiesis in Multiple Myeloma
期刊:Nature Communications
发表时间:2023 年
影响因子:15.7
在多发性骨髓瘤中,血小板减少是常见并发症之一,但其背后的代谢机制长期并不清楚。在 《Nature Communications》发表的研究发现,骨髓微环境中过量丝氨酸会损害巨核细胞生成和血小板生成。
研究团队通过非靶向代谢组学发现,多发性骨髓瘤患者骨髓微环境中丝氨酸水平显著升高。随后进一步结合 [U-¹³C₃]-Serine 和 [U-¹³C₂]-Glycine 稳定同位素示踪,追踪丝氨酸进入一碳代谢、甲硫氨酸循环以及相关表观遗传调控过程。
结果表明,过量丝氨酸可通过影响一碳代谢和甲硫氨酸循环,调控 H3K9 三甲基化等表观遗传修饰,进而抑制巨核细胞和血小板生成。
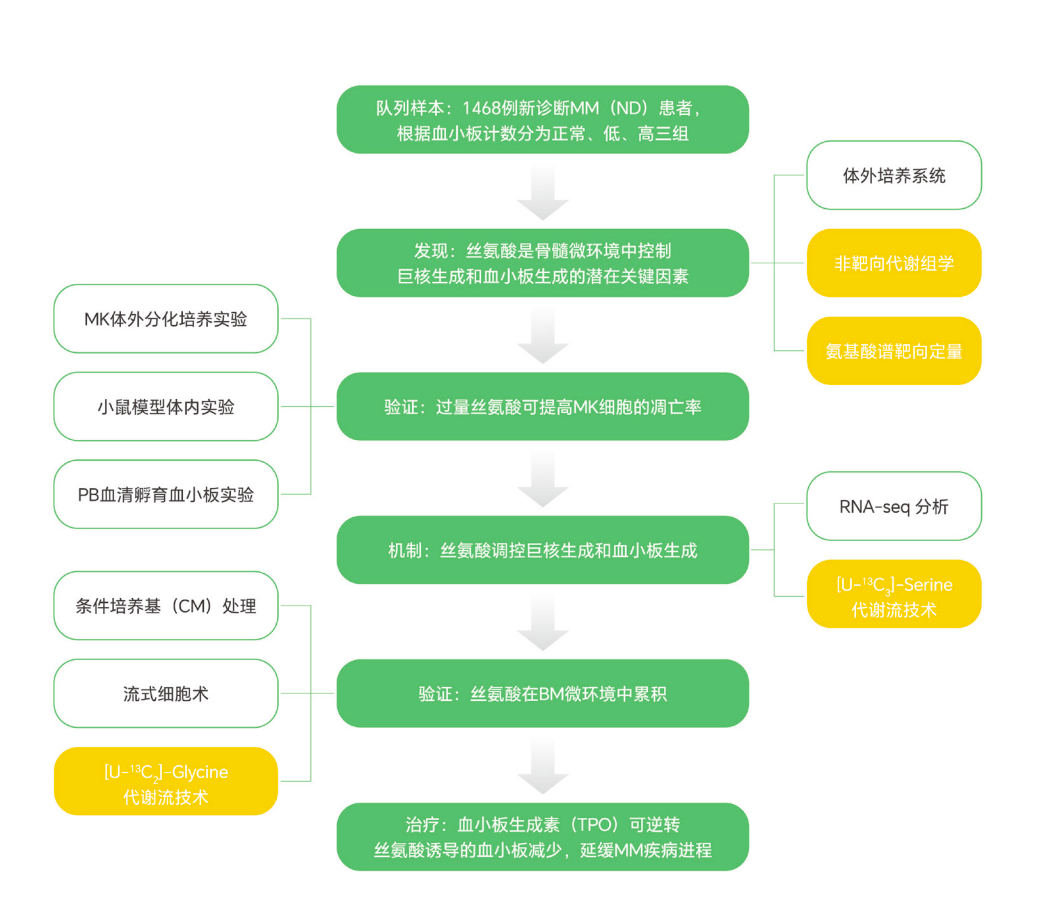
这个案例很好地体现了代谢流分析的价值:
它不仅能发现“哪个代谢物变了”,更能进一步回答这个代谢物从哪里来、流向哪里、以及如何改变细胞命运。
八、绘云生物:从代谢流到多组学的一站式科研解决方案
代谢流分析让我们能够从“代谢物变化”进一步走向“代谢机制解析”。
但在真实科研中,一个生物学问题往往并不只涉及代谢层面。
例如,肿瘤耐药可能同时涉及代谢重编程、蛋白修饰、转录调控和微环境变化;免疫细胞功能改变,可能既有能量代谢变化,也伴随细胞因子、膜蛋白、信号通路和菌群代谢物的共同调控。
因此,越来越多研究已经从单一组学,走向多组学联合解析。
绘云生物长期专注于医学代谢组学、多组学技术和稳定同位素代谢流平台建设,致力于为科研、临床和产业客户提供从实验设计、样本检测、数据分析到机制解释的一站式解决方案。
同位素示踪代谢流部分检测列表

同位素示踪代谢流样本用量

为什么选择绘云生物?
绘云生物不仅提供检测服务,更关注科研问题本身。
从研究方案设计、实验路线优化、样本处理建议,到质谱检测、数据分析和机制解释,绘云生物希望帮助客户真正回答:
* 哪条通路发生了变化?
* 哪个代谢物可能是关键节点?
* 代谢变化背后的机制是什么?
* 如何进一步设计验证实验?
* 如何将组学结果转化为文章中的机制故事?
截至目前,绘云生物已协助客户发表 SCI 论文 400+ 篇,累计影响因子 4000+,平均影响因子 >10,相关成果发表于 Cell、Science、Nature、Cancer Cell、Cell Metabolism、Nature Metabolism、Immunity、Gut、PNAS 等国际期刊。
欢迎交流合作
如果您的课题涉及:
代谢组学、稳定同位素代谢流、蛋白组学、蛋白修饰组学、转录组学、宏基因组学、多组学联合分析,或希望从代谢变化进一步深入机制解析,欢迎联系绘云生物。
绘云生物将为您提供从实验设计到数据解析的一站式科研支持。
联系方式:
电话:18516553756
官网:[www.hmibiotech.com](http://www.hmibiotech.com)
邮箱:[linshitong@hmibiotech.com](mailto:linshitong@hmibiotech.com)
二维码:

绘云生物公众号

项目联系










还没有人评论,赶快抢个沙发