 腾讯登录
腾讯登录你会被饿着,你的DNA呢?
| 导读 | 父母的营养状况会对其生殖细胞的表观遗传学修饰带来影响,进而影响到后代的健康状况。 |
父母的营养状况会对其生殖细胞的表观遗传学修饰带来影响,进而影响到后代的健康状况。
人类研究和动物研究都表明,在出生前,即宫内时所处的大环境会影响到子代成年之后的身体健康状况及患病情况。流行病学调查也发现,母亲在妊娠时如果遭遇饥饿,或者父系祖辈营养过剩,都会增加子代患上心血管疾病和糖尿病的风险。不论是饥饿还是营养过剩,都会给身体健康带来不良的代谢反应,而且这种不良的反应还会一代代地传下去。同样的,如果用低蛋白饮食喂养怀孕的大鼠,那么连续两代子代大鼠在成年之后都会患上糖尿病。虽然有这么多研究都表明,在宫内时受到的各种环境因素影响都会给子代成年之后,以及这些子代的后代带来身体健康方面的影响,但是这种多代遗传的机制还是没搞清楚。
Radford等人在这方面开展的新工作,他们用一种全基因组深度分析技术对营养状况不佳的小鼠动物模型开展了研究。这种小鼠动物模型是一种低出生体重动物模型,其子代和孙代都存在糖耐量异常(glucose intolerance)和胰腺功能减低的问题。Radford等人不仅向我们展示了足够的证据,来解释这种多代遗传的作用机制,同时还为我们指明了一条研究方向,能够在这个领域更加深入地发掘下去。
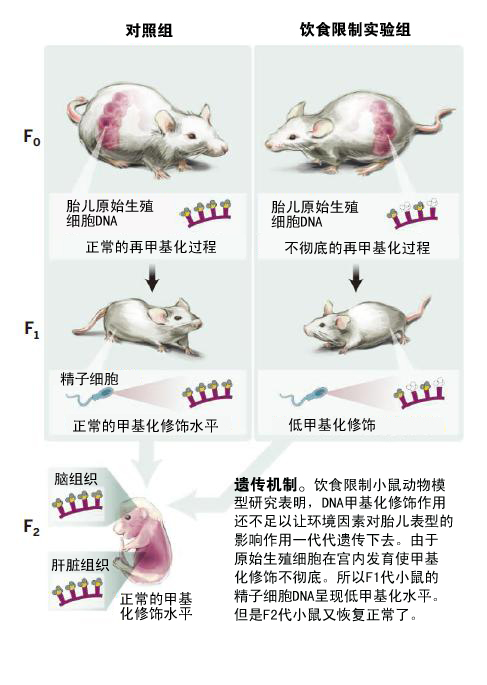
由于这种宫内暴露因素对子代的影响作用都显现得比较晚(通常都在子代成年之后才会表现出来),所以大家通常都认为是表观遗传学机制(Epigenetic mechanisms)在其中起到了主要的作用。即并非由于DNA序列发生改变才带来了这种可遗传的表型改变。表观遗传学机制主要包括DNA甲基化修饰(DNA methylation)、染色质结构(chromatin structure)改变、非编码RNA(noncoding RNA)改变以及核组织(nuclear organization)改变等。而涉及表型遗传最多见的就是DNA甲基化修饰作用。其他一些宫内暴露动物模型研究已经发现,人类IGF2基因和大鼠PPARα基因的甲基化修饰发生改变就与宫内暴露因素遗传作用相关。由此可见,如果缺乏其他的环境刺激,那么宫内影响因素能够继续遗传下去的原因可能就是通过生殖细胞的表观遗传学遗传机制(germline epigenetic inheritance)了。即在宫内发生了重编程的(reprogrammed)胎儿的精子细胞或者卵子细胞的表观遗传学信号被遗传下去了。
为了检验这个猜想(DNA甲基化修饰水平发生改变会影响营养水平不佳小鼠的子代表型)是否正确,Radford等人利用甲基化DNA免疫沉淀测序(methylated DNA immunoprecipitation–sequencing, MeDIP–sequencing)及重亚硫酸盐焦磷酸测序技术(bisulfite pyrosequencing)进行了研究,结果发现小鼠宫内卡路里不足会影响子代(F1代)雄性小鼠精子细胞的位点特异性DNA甲基化修饰水平(locus-specific DNA methylation patterns),如图所示。这种营养状况不良的情况主要发生在孕晚期,此时初级精子细胞已经完成了表观遗传学重编程过程,形成了雄性生殖细胞特有的DNA甲基化修饰。科研人员们选择了四窝代表性的小鼠幼崽进行了对照试验。在营养状况不佳的F1代雄性小鼠的基因组中一共发现了111个低甲基化修饰的区域(hypomethylated regions)。从中随机选择了24个区域进行试验验证,结果有17个区域都得到了确认,这说明有70%的低甲基化区域真的受到了宫内营养状况不良的影响。虽然也发现了一些超甲基化区域(hypermethylated regions),但是用重亚硫酸盐焦磷酸测序试验却没有得到验证,这也提示我们,针对不同的样品,需要用不同的试验方法进行验证。
Radford等人为了验证这些表观遗传学修饰情况已经发生了改变的F1代小鼠是否能够将这种改变继续遗传下去,又将处于糖尿病前期的F1代雄性幼鼠与对照组雌性幼鼠杂交,然后对胚胎龄为16.5天的F2代小鼠的肝脏和大脑进行了DNA甲基化水平分析。他们之所以选择这种父系遗传策略(paternal transmission strategy)就是为了去除母亲在孕期带来的一些干扰。通过对胚胎晚期F2子代组织的分析,发现之前处于低甲基化水平的DNA序列的甲基化水平又得到了恢复,在对照组和饮食受限的F2子代小鼠之间已经没有区别了。不过有一些紧邻那些低甲基化区域的基因的表达水平还是有所区别。所有这些试验结果表明,DNA甲基化机制可能不是孕期环境因素能够持续遗传下去(遗传给F2代)的主要表观遗传学作用机制,不过这也需要进一步验证。不过该试验结果与另外一个同样使用了饮食限制方法的小鼠动物试验得到的结果有一点出入,该试验在F2子代小鼠中发现,Lxra基因的甲基化修饰水平还是有所差异。不过该试验采用的试验方法有所不同,他们的研究对象是转录水平的差异,首先在对照组和饮食限制试验组之间寻找基因在转录水平的差异,然后再对相关序列进行分析。当然,这两组试验在饲料和饲养方法上的差异也有可能带来试验结果上的不一致。
孕期营养不良对胎儿的影响是如何一代代遗传下去的,其作用机制目前还不清楚。在F1代精子细胞基因组中发现的低甲基化区域表明,DNA甲基化修饰作用介导了环境干扰因素对发育的改变,但是一定还有其他的表观遗传学机制在其中发挥了作用。这种MeDIP测序方法没能发现的其他DNA甲基化修饰异常位点也有可能与子代发育有关。亦或有其他的表观遗传学修饰机制在这些位点发挥了作用,将环境干扰因素对发育的改变作用一代代的遗传了下去。在精子细胞富含核小体的发育位点中发现了组蛋白H3 Lys4和组蛋白H3Lys27的三甲基化修饰(分别生成H3K27me3和H3K4me3)现象,这说明组蛋白修饰可能是一种遗传作用机制。另外一种可能的作用机制就是小RNA(small RNA)作用机制,正如我们在秀丽隐杆线虫饮食限制模型(Caenorhabditis eleganscaloric restriction model)中观察到的那样。
Radford等人为我们提供了一个范例,可以先用全基因组研究技术缩小研发范围,然后再一一加以验证,今后在类似的工作中也都可以使用这种策略。虽然DNA甲基化修饰作用在营养缺乏所导致的发育改变过程中起到了重要的作用,但是其他的表观遗传学作用机制在这种因为环境因素所带来的遗传效应中的作用也不应该被忽视。

{kind=link}
还没有人评论,赶快抢个沙发